Deriva genética
Parte de la serie de |
Evolución biológica |
|---|
 |
Temas clave Biología evolutiva Introducción a la evolución Antepasado común Evidencia de antepasado común |
Historia de la idea Información general Renacimiento e Ilustración Antes de Darwin Darwin • Su libro Antes de la Síntesis Síntesis evolutiva moderna Evolución molecular • Evo-devo |
Historia Natural Historia de la vida Biodiversidad • Biogeografia Clasificación • Cladística Paleontología •Filogenia Evolución humana |
Procesos y Resultados Genética de poblaciones Variación Mutación Selección natural • Adaptación Deriva genética • Flujo genético Especiación • Radiación adaptativa Cooperación • Coevolución Extinción |
Implicaciones Sociales Teoría y hechos Efectos sociales Controversias Objeciones Nivel de aceptación |
Campos y aplicaciones Aplicaciones de la evolución Selección artificial Bioinformática Antropología evolucionista Computación evolutiva Evolución experimental Genética ecológica Neurociencia evolucionista Fisiología evolucionista Psicología evolucionista Sistemática |
Portal de biología Categoría |
La deriva genética, también conocida como la deriva génica, la deriva alélica o efecto Sewall Wright[1] (por el biólogo Sewall Wright), es una fuerza evolutiva que actúa junto con la selección natural cambiando las frecuencias alélicas de las especies en el tiempo. Es un efecto estocástico que es consecuencia del muestreo aleatorio en la reproducción y de la pérdida de unos alelos por azar y no por selección natural. Se trata de un cambio aleatorio en la frecuencia de alelos de una generación a otra.[2] Normalmente se da una pérdida de los alelos menos frecuentes y una fijación (frecuencia próxima al 100%) de los más frecuentes, resultando una disminución en la diversidad genética de la población.
Al igual que la selección natural, actúa sobre las poblaciones, alterando la frecuencia de los alelos (frecuencia alélica) y la predominancia de los caracteres sobre los miembros de una población, y cambiando la diversidad genética del grupo. Los efectos de la deriva se acentúan en poblaciones de tamaño pequeño (como puede ocurrir en el efecto de cuello de botella o el efecto fundador), y resultan en cambios que no son necesariamente adaptativos.
La deriva genética tiende a formar una población homocigótica, es decir tiende a eliminar los genotipos heterocigóticos. Además, ya que en cada población pueden ser distintos los alelos que se pierden y se fijan, la deriva hace que dos o más poblaciones de la misma especie tiendan a diferenciarse genéticamente.
La frecuencia de un gen puede cambiar de una generación a otra gracias a lo que se llaman errores de muestreo, ya que de todos los genes de la población sólo una pequeñísima fracción pasará a la siguiente.
El error de muestreo se produce cuando los gametos se unen para producir la progenie. Muchos organismos producen un número grande de gametos pero, cuando el tamaño de la población es pequeño, un número limitado de gametos se une para producir los individuos de la generación siguiente. El azar influye en que alelos están presentes en esta muestra limitada y, de esta manera, el error de muestreo puede conducir a la deriva genética o cambios en las frecuencias alélicas. Dado que las desviaciones de las proporciones esperadas son aleatorias la dirección del cambio es imprevisible. No obstante, podemos predecir la magnitud de los cambios.
Índice
1 Magnitud de la deriva genética
2 Tamaño efectivo de una población
3 Causas de la deriva genética
3.1 Recursos
3.2 Efecto fundador
3.3 Cuello de botella
4 Efectos de la deriva genética
5 Conclusión
6 Ejemplos
7 Véase también
8 Referencias
9 Bibliografía
Magnitud de la deriva genética
La cantidad de deriva genética puede estimarse a partir de la varianza en la frecuencia alélica. Suponga que observamos un número grande de poblaciones separadas, cada una con individuos N y frecuencias alélicas de p y q. Después de una generación de apareamiento al azar la deriva genética expresada en términos de la varianza en la frecuencia alélicas entre las poblaciones (sp2) será:
sp2=pq2N{displaystyle sp2={pq over 2N}}

La cantidad de cambio que resulta de la deriva genética es determinada por dos parámetros: las frecuencias alélicas (p y q) y el tamaño de la población (N). La deriva genética será máxima cuando p y q sean iguales.
Ejemplo: suponga que una población consiste en 50 individuos. Cuando las frecuencias alélicas sean iguales (p = q = 0,5), la varianza en la frecuencia alélica (sp2) será (0,5 . 0,5)/(2 . 50) = 0,0025. En contraposición, cuando p = 0,9 y q = 0,1, la varianza en la frecuencia alélicas sólo será de 0,0009. Así mismo la deriva genética será más alta cuando el tamaño de la población sea pequeño. Si p =q = 0,5, pero el tamaño de la población es sólo 10 en lugar de 50, entonces la varianza en la frecuencia alélicas se vuelve de (0,5 . 0,5)/(2 . 10) = 0,0125, lo que es cinco veces mayor que cuando el tamaño de la población era de 50.
Esta divergencia de poblaciones mediante la deriva genética se ilustra bien con los resultados de un experimento llevado a cabo por Peter Buri en la mosca de la fruta. Buri examinó la frecuencia de dos alelos (bw75 y bw), ambos con frecuencia 0,5, que afectan al color de ojos de Drosophila en 107 poblaciones replicadas en 19 generaciones. El resultado de la deriva genética fue que las frecuencias alélicas en poblaciones diferentes divergieron y a menudo se volvieron fijas para un alelo o para el otro.
Tamaño efectivo de una población
Cuanto mayor sea el número de individuos de la población, menor será la diferencia entre las frecuencias de una generación y otra, aunque lo que cuenta no es el número real de individuos, sino lo que se llama tamaño eficaz. El tamaño eficaz o efectivo de una población se define por aquellos individuos que dejan descendientes, que en el caso de casi todos los organismos puede ser un número mucho menor que el total de individuos, ya que sólo son los individuos reproductores aquellos que trasmiten sus genes.
Ese tamaño efectivo se calcula con una fórmula matemática:
Ne=(4.nomachos.nohembras)(nomachos+nohembras){displaystyle N_{e}={frac {(4.n^{o}machos.n^{o}hembras)}{(n^{o}machos+n^{o}hembras)}}}


Por ejemplo, cuando una población consiste en 90 machos y 10 hembras, el tamaño efectivo de la población es de sólo 36 y la deriva genética sucederá como si la población real consistiera de sólo 36 individuos. De igual forma si fuesen 10 machos y 90 hembras.
Otros factores que influyen en el tamaño efectivo de la población incluyen la variación entre los individuos en el éxito reproductivo, las variaciones en el tamaño de la población y si el apareamiento es aleatorio.
También hay que decir que el tamaño efectivo de la población disminuye cuando hay cantidades desiguales de machos y hembras con capacidad reproductora.
Causas de la deriva genética
Si bien todas las derivas genéticas se originan a partir del error de muestreo, existen varias maneras diferentes por las que puede originarse dicho error.
Recursos
Una población puede verse reducida en tamaño durante varias generaciones debido a limitaciones en el espacio, los alimentos o algún otro recurso crítico.
Efecto fundador
Este se debe a la instalación de una población por un número pequeño de individuos. Aunque una población puede aumentar y volverse bastante grande, los genes portados por todos sus miembros derivan de los pocos genes presentes originalmente en los fundadores (considerando que no hay migración ni mutación). Los acontecimientos al azar que afectan algunos genes presentes en los fundadores tendrán una influencia importante en la composición de la población general.
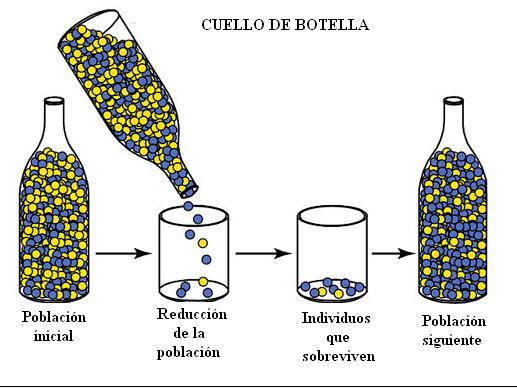
Cuello de botella
Consiste en que una población sufre una reducción drástica en su tamaño, dando esto lugar a una población con alta probabilidad de sufrir deriva genética. Cualquier mutación en un individuo se amplificará en la comunidad cuando esta crezca de nuevo. Los eventos de cuello de botella también son caracterizados por producir una reducción de la diversidad genética de las especies, debido a la extinción de los alelos menos frecuentes.
Efectos de la deriva genética
La deriva genética tiene varios efectos importantes en la composición genética de una población.
1.- Produce un cambio en las frecuencias alélicas dentro la población. Dado que la deriva es aleatoria, es tan frecuente que las frecuencias alélicas aumenten como que disminuyan y puede deambular con el paso del tiempo.[3]
2.- Reduce la variación genética dentro de las poblaciones. Llegando incluso a que un alelo alcance la frecuencia de 1, se dice que el alelo se ha fijado, o cero, cuando el alelo desaparece por completo en la población.
Conclusión
Si no hubiera otros procesos de cambio evolutivo, tales como la mutación y la selección natural, las poblaciones llegarán al final a tener un solo alelo de cada gen, aunque se tardase muchas generaciones en llegar a ello. La razón es que, tarde o temprano, uno u otro alelo sería eliminado por la deriva genética sin posibilidad de que reapareciera por mutación o migración. Debido a la mutación de alelos desaparecidos de una población puede volver a reaparecer de nuevo, y gracias a la selección natural, la deriva genética no tiene consecuencias importantes en la evolución de especies, excepto en poblaciones de pocos individuos.

Gráfica de simulación en la fijación de alelos.
Ejemplos
La deriva genética es un proceso que ha afectado a gran cantidad de poblaciones de diferentes especies a lo largo del tiempo. Algunos de esos casos son:
El elefante marino del norte sufrió un cuello de botella genético intenso entre 1820 y 1880, con un proceso de deriva genética a continuación. En la actualidad estas focas tienen niveles bajos de variación genética.[4]
Al igual le pasa a la gaviota argéntea en cuya especie se puede observar una deriva genética incluso en una misma época en lugares diferentes. La gaviota argéntea que se puede ver en Cádiz presenta manchitas, y sin embargo según vamos hacia el oeste dándole la vuelta al planeta se pueden ir observando las distintas variedades de este tipo de gaviota. Por ejemplo en América es totalmente blanca; en Asia ya comienza a ser moteada y de ese modo cada vez son más las manchitas hacia la Europa del Este, hasta que llegamos a Cádiz y encontramos a ambas poblaciones: una completamente moteada y otra blanca, como si fuesen dos especies distintas. Son los extremos de una misma especie con deriva genética y geográfica que no se aparean entre sí.

Fijación de alelos.
Otro ejemplo es el de la tortuga Testudo hermanni, también con una población muy homogénea genéticamente debido a un cuello de botella que en algún momento sufrió la población seguido de un proceso de deriva genética.
Véase también
- Motoo Kimura
- Teoría neutralista de la evolución molecular
Referencias
↑ The Structure of Evolutionary Theory (2002) por Stephen Jay Gould, Capitulo 7, sección "Synthesis as Hardening"
↑ Pierce, Benjamin A. (2009-07). Genética: Un enfoque conceptual. Ed. Médica Panamericana. ISBN 9788498352160. Consultado el 9 de diciembre de 2017.
↑ Pierce, Benjamin A. (2009-07). Genética: Un enfoque conceptual. Ed. Médica Panamericana. ISBN 9788498352160. Consultado el 9 de diciembre de 2017.
↑ Audesirk, Teresa; Audesirk, Gerald; Byers, Bruce E. (2003). Biología: la vida en la tierra. Pearson Educación. ISBN 9789702603702. Consultado el 9 de diciembre de 2017.
Bibliografía
- Genética: Un enfoque conceptual. Pierce, Benjamin A. Editorial Médica Panamericana, S.A., 2ª edición. ISBN 978-84-7903-889-2, ISBN 84-7903-889-6
- Conceptos de genética. Klug William S. Ed. Pearson, ISBN 978-84-205-5014-5, ISBN 84-205-5014-0


